
水草在面對水中 CO2 不足的環境中,
必須想辦法來攝取有機溶解無機碳(DIC),
C-4 光合作用是其中一種方式,
讓我們一起來瞭解一下何謂 C-4 光合作用。
從陸生植物的角度來看,
C4 植物主要發生於熱帶或亞熱帶的植物,
例如玉米、甘蔗、高樑、狗尾草、馬齒莧...等等。
C4 植物比 C3 植物具有較強的光合作用,
從生理上來看,
C4 植物內的磷酸烯醇丙酮酸羧化酶(PEPC)固定 CO2 的活性較強以及光呼吸很弱有關,
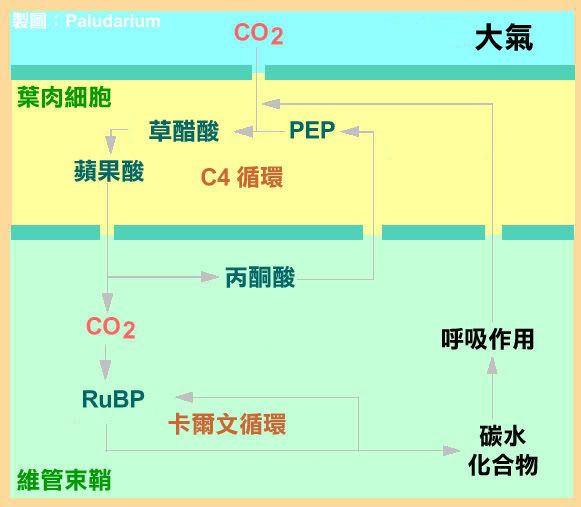
C4 途徑的 CO2 固定最初是由 PEPC 來完成的,
PEPC 對 HCO3 的 Km 值是 7 μM,
而核酮糖二磷酸縮化酵素(rubisco)對 CO2 的 Km 值是 450 μM,
代表著前者對 CO2 的親和力比後者大很多。
因此 C4 植物的 CO2 固定速率比 C3 植物高,
所以陸生 C4 植物的 CO2 補償點比較低(0-5 ppm),
而陸生 C3 植物的 CO2 補償點介於 20-50 ppm 之間。
這有個好處是,
當外界乾旱導致植物氣孔部分關閉時,
C4 植物仍能利用低含量的 CO2 繼續生長,
但 C3 植物就不能了。
所以在乾旱的環境中,
C4 植物生長比 C3 植物來得好。
「陸生」植物三種光合作用的比較,此表僅供參考,不可直接套用至「水草」。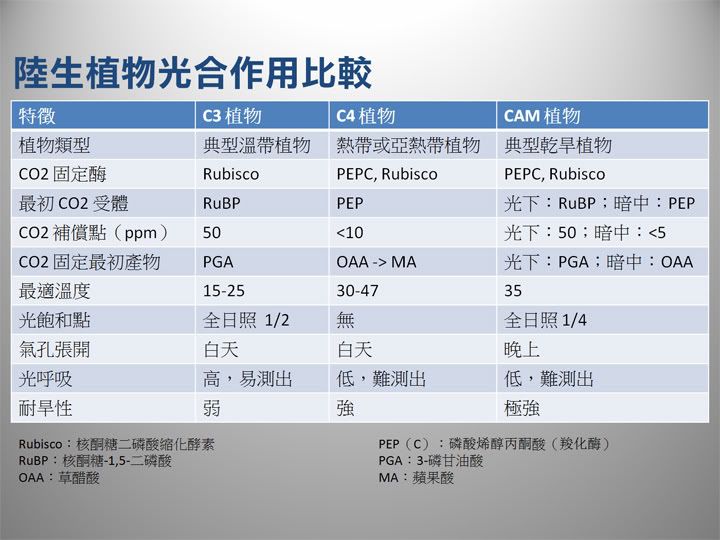
但在水中環境並沒有乾旱的問題,
在大多數的情況下,
這是對周遭 CO2 濃度低下時的反應,
因為在 CO2 充足的環境下,
這些水草又會變回 C-3 方式的碳固定。
具有這種行為模式且最被人研究最透徹的,
要算是黑藻(Hydrilla verticillata)了。
黑藻在夏季常日照、高溫且低 CO2 環境下,
會轉變成 C-4 酸合成方式。
Keeley 於 1990 年指出,
被認為會進行 C-4 酸合成的所有水草,
其實在體內的每一個細胞當中,
C-4 的碳固定是和 C-3 的碳固定同時進行的。
除了黑藻以外,
C-4 水草的種類還包括了水蘭(Vallisneria spiralis)、牛毛氈(Eleocharis acicularis)、E. baldwinii 和 Orcuttia californica。
黑藻是最常被拿來研究的水草之一,由於同時兼具 C3 與 C4 的能力,生命力極強,是著名的水生雜草。

根據水草 C-4 碳固定的模式來看,
兩種羧化酶(carboxylase)酵素~ PEPC 與 rubisco ~分別位於光合作用細胞內的不同位置上,
而不是像陸生植物的 C-4 植物一樣位於不同的細胞內。
PEPC 乃是位於細胞質內,
而 rubisco 則位於葉綠體內。
當 CO2 擴散至細胞內,
會被 PEPC 或 rubisco 兩者之一給固定。
蘋果酸(Malate)或 C-4 合成的其他有機酸產物,
會由細胞質運送至葉綠體內,
並且在此處會去羧基(decarboxylate)變成 CO2,
而交由 rubisco 再固定,
隨後進入卡爾文循環中的 3-磷甘油酸(3-PGA)部分。
這樣的結果使得 rubisco 的 CO2 濃度上升,
減少了光呼吸作用。
在 C-4 水草的體內,
並沒有如陸生 C-4 植物一樣的花環型結構(Kranz type)。
Keeley 於 1990 年描述了挺水的水草 Orcuttia californica,
此草由水中葉發展成浮水葉乃至於水上葉。
其浮水葉與水上葉都具備典型的花環型結構,
而其水中幼葉缺乏這項結構,
然而其 C-4 固定的產量與陸生 C-4 植物卻是相當的。
水草雖然也具有 C4 光合作用的模式,
但不論在結構上或發生部位上,
甚至在發生的環境上,
都與陸生植物不盡相同。
這就是我們一再強調的:
不可將陸生植物的研究原封不動的套用至水草生理上。
本文同時刊載於台灣「AquaPets」雙月刊 2008 年八月號。

 RSS 訂閱
RSS 訂閱