

Tom Barr 與水草導師團隊
作者:Tom Barr
翻譯:Erich Sia
水草要努力應付二氧化碳在每小時、每天和每周之間的改變,
也要花更多的能量來適應二氧化碳,
然後成長、獲得營養和捕捉光線等等。
在無二氧化碳的系統中,
經常換水導致每周之間的二氧化碳濃度的激增,
好幾個月才換一次水會比較好。
對二氧化碳豐富(不受限制系統)的缸子來說,
換水就沒這樣的衝擊。
水草不需要適應二氧化碳,
因為水草本來就有充裕的二氧化碳,
而且任何時候都在供應。
翻譯:Erich Sia
水草要努力應付二氧化碳在每小時、每天和每周之間的改變,
也要花更多的能量來適應二氧化碳,
然後成長、獲得營養和捕捉光線等等。
在無二氧化碳的系統中,
經常換水導致每周之間的二氧化碳濃度的激增,
好幾個月才換一次水會比較好。
對二氧化碳豐富(不受限制系統)的缸子來說,
換水就沒這樣的衝擊。
水草不需要適應二氧化碳,
因為水草本來就有充裕的二氧化碳,
而且任何時候都在供應。
我們也可以略加限制二氧化碳和磷酸,
並且擁有還算穩定的系統,
但這些比較不易複製。
儘管如此,
只要在二氧化碳夠穩定來維持組織內相同的 Rubisco 濃度,水草終將能夠適應中濃度的二氧化碳。
所以通常在低二氧化碳系統中不進行或減少換水,
那麼就像窮人的施肥滴液法所建議的 10-15 ppm 二氧化碳濃度,
效果也不錯且能減少藻類。
儘管如此,
這些系統還是限制了二氧化碳,
水草已經適應了,
就像在無二氧化碳系統一樣。
只要二氧化碳的濃度穩定,
並且在每周之間的變動不大,
水草應該還能長得不錯且長得慢,
不過也沒藻類。
並且擁有還算穩定的系統,
但這些比較不易複製。
儘管如此,
只要在二氧化碳夠穩定來維持組織內相同的 Rubisco 濃度,水草終將能夠適應中濃度的二氧化碳。
所以通常在低二氧化碳系統中不進行或減少換水,
那麼就像窮人的施肥滴液法所建議的 10-15 ppm 二氧化碳濃度,
效果也不錯且能減少藻類。
儘管如此,
這些系統還是限制了二氧化碳,
水草已經適應了,
就像在無二氧化碳系統一樣。
只要二氧化碳的濃度穩定,
並且在每周之間的變動不大,
水草應該還能長得不錯且長得慢,
不過也沒藻類。
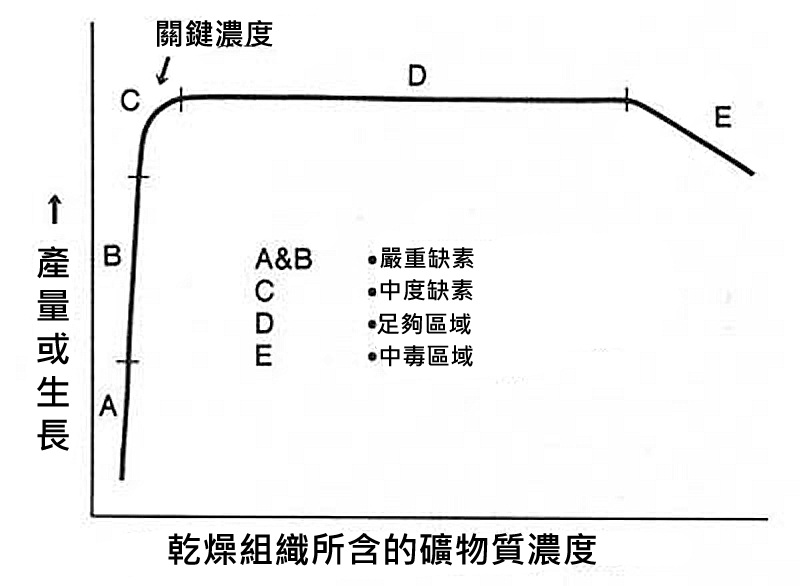
下面圖表的曲線顯示:
在較低濃度下,
在限制的範圍內產量減少了。
這並非黑白分明的區別,
而是有各種程度不同的限制。
輕度的限制對整體成長的影響較小,
但對二氧化碳的影響就很大了。
非常嚴重的限制對於成長和缺素就有很顯著的影響。
在差強人意和輕度限制的區域,
留給水族愛好者在施肥時最大的餘地。
不論是二氧化碳、營養或照明。
唯有二氧化碳(營養或照明)在充足的區域時,
我們才能確保我們在測試其他參數時,
彼此之間是相互獨立的。
在較低濃度下,
在限制的範圍內產量減少了。
這並非黑白分明的區別,
而是有各種程度不同的限制。
輕度的限制對整體成長的影響較小,
但對二氧化碳的影響就很大了。
非常嚴重的限制對於成長和缺素就有很顯著的影響。
在差強人意和輕度限制的區域,
留給水族愛好者在施肥時最大的餘地。
不論是二氧化碳、營養或照明。
唯有二氧化碳(營養或照明)在充足的區域時,
我們才能確保我們在測試其他參數時,
彼此之間是相互獨立的。
回到藻類的問題。
營養問題是否可能扮演了某個角色?
我用氨氮(NH4)來試驗,
發現在高劑量(1-2 ppm)且強光環境下,
會誘發綠水。
降低二氧化碳也會促進綠水的長速率。
綠水在新設置的強光水族缸很常見,
這些缸子尚未進行氮循環,
意思是說缸內的細菌數量不足且水草生長不良,
無法將氨氮循環成硝酸(NO3)。
氨氮是我發現唯一能夠誘發綠水的營養,
同樣的道理,
水族缸內日漸增加的生物負荷,
到了一個臨界點以後就會爆藻,
效果是類似的。
這說明了氨氮扮演著某個角色。
然而其他人和我一樣投予氨氮,
可並無法誘發綠水或其他的藻類。
所以這些結果似乎並無定論。
或許不良的二氧化碳和照明強度扮演著一定的角色?
或許泥炭苔、鞣酸、沉積質的種類也都扮演著一定的角色?
有些人也持相同的看法。
不過,
二氧化碳(低於無限制濃度下的變化)和強光卻是非常一致的。
這兩者能夠誘發許多藻類品種。
低磷酸似乎誘發了綠斑藻。
低二氧化碳則誘發了黑毛藻和鹿角藻(在高生物負荷量下),
藍綠藻似乎與低硝酸、有機質有關。
每一種藻類都有自己的一套關聯變數,
也可能爆藻的因素不只一個。
儘管如此,
在另一個獨立的系統內,
透過每次只操控一個變因的方式來測試藻類的誘發,
是了解藻類為何成長以及如何加以阻止的最好方法。
受控制的不平衡法(MCI)建議極度限制磷酸達幾個星期,
直到出現綠斑藻為止,
然後再把磷酸添加回來,
而當其他藻類都消失了,
隨後也能擺脫綠斑藻。
在爆綠水的情況下,
其他的藻類也鮮有繼續生長的,
所以我們能夠用藻類和限制磷酸等等做法,
或者也可費力的控制誘因來消滅藻類。
二氧化碳(低於無限制濃度下的變化)和強光卻是非常一致的。
這兩者能夠誘發許多藻類品種。
低磷酸似乎誘發了綠斑藻。
低二氧化碳則誘發了黑毛藻和鹿角藻(在高生物負荷量下),
藍綠藻似乎與低硝酸、有機質有關。
每一種藻類都有自己的一套關聯變數,
也可能爆藻的因素不只一個。
儘管如此,
在另一個獨立的系統內,
透過每次只操控一個變因的方式來測試藻類的誘發,
是了解藻類為何成長以及如何加以阻止的最好方法。
受控制的不平衡法(MCI)建議極度限制磷酸達幾個星期,
直到出現綠斑藻為止,
然後再把磷酸添加回來,
而當其他藻類都消失了,
隨後也能擺脫綠斑藻。
在爆綠水的情況下,
其他的藻類也鮮有繼續生長的,
所以我們能夠用藻類和限制磷酸等等做法,
或者也可費力的控制誘因來消滅藻類。

儘管如此,
我們能夠透過降低照明或者搭配良好的二氧化碳穩定度,
同樣的來消除藻類。
一般而言,
水族愛好者發現到,
留意照明和二氧化碳(當然還有水草),
更好用也更成功。
營養的投予或添加其實相當的容易,
水草會適應高濃度或低濃度,
就像二氧化碳一樣,
只要供給維持穩定的話。
很多人大錯特錯的是他們的結論,
認為更多的營養會誘發藻類。
這完全是不正確的。
這只發生在附屬於其他變因之時,
例如二氧化碳。
雖然這個說法有可能會激怒推行這種限制磷酸方式的人......
當他們在被告知並非因為限制方式才造成的,
這直接挑戰了他們的主張。
事實就是如此,
那些都是結果,
而沒辦法調解的是那個假說:
限制磷酸才能控制藻類。
這是能夠檢驗況且也已經被證明是虛假的。
他們的假說沒法解釋高濃度磷酸怎麼無法誘發藻類。
如果假說認為高磷酸濃度就等於藻類,
那這個假說就必須接受檢驗。
假說可以任何的方式去改寫或重新包裝,
但就是沒個別強調照明、二氧化碳和營養,
其方法仍舊會令很多人獲得錯誤的結論。
這並不暗指限制磷酸並非藻類的控制的可行選項,
限制磷酸的方法已經幾十年了,
但是其作用被曲解成了藻類方面的限制。
想得到藻類萌芽的根本原因,
要在控制選擇上採取產值更高的方法。
我們現在有更多的工具來達到藻類的控制,
並不只有限制磷酸而已,
也包括了諸如增加二氧化碳、降低照明強度、較好的循環、更好的過濾器清理、維持固定的生物負荷等等。
搭配液態碳元素的添加和換水(增加或減少),
此外還有無二氧化碳法,
調和了為什麼不同的方法也可以有效地促成整體的水草生長。
這也容許我們進行更好的測試,
並找出更多的原因,
而不是把我們所有的「雞蛋」都放在「營養」的這個籃子裡。
那樣實在太危險了。
他們的假說沒法解釋高濃度磷酸怎麼無法誘發藻類。
如果假說認為高磷酸濃度就等於藻類,
那這個假說就必須接受檢驗。
假說可以任何的方式去改寫或重新包裝,
但就是沒個別強調照明、二氧化碳和營養,
其方法仍舊會令很多人獲得錯誤的結論。
這並不暗指限制磷酸並非藻類的控制的可行選項,
限制磷酸的方法已經幾十年了,
但是其作用被曲解成了藻類方面的限制。
想得到藻類萌芽的根本原因,
要在控制選擇上採取產值更高的方法。
我們現在有更多的工具來達到藻類的控制,
並不只有限制磷酸而已,
也包括了諸如增加二氧化碳、降低照明強度、較好的循環、更好的過濾器清理、維持固定的生物負荷等等。
搭配液態碳元素的添加和換水(增加或減少),
此外還有無二氧化碳法,
調和了為什麼不同的方法也可以有效地促成整體的水草生長。
這也容許我們進行更好的測試,
並找出更多的原因,
而不是把我們所有的「雞蛋」都放在「營養」的這個籃子裡。
那樣實在太危險了。

就手段方面而言,
我們需要一個假說,
這個假說強調了多種的方法、多種似乎相違背的觀察,
以及解釋為何水族愛好者所選定每種方法能夠有效。
這個假說給了我們最好的解釋、最多的工具,
而且也最合理。
這個假說也有助於我們把焦點放在水草的成長,
況且藻類並不會跟著出現,
也讓我們知道每個藻類品種是哪一種指標生物或"測試工具"。
這同時讓控制藻類和促進水草生長在管理上容易多了。
再來我們看看雷德菲爾德比例(Redfield ratio)。
下述是 Anderson 這位專家在 2005 年時針對「藻類栽培方法」所發表的一段看法:
「雷德菲爾德比例(RR)可說是是水族生態領域最常被濫用的參數之一,
雷德菲爾德比例是個近似值,
由非常大規模的時間和空間的平均組合而成,
並沒有去敘述個別細胞和族群。」
Anderson 繼續引述幾個同相(Chemostat)研究,
舉出淡水柵藻(Scenedesmus)的氮磷比(N:P)是 30:1,
幾乎是兩倍的差異,
而巴夫藻(矽藻)(Pavlova)的氮磷比是 25 至 33,
角刺藻(矽藻)(Chaetoceros)的氮磷比則是 35-40。
這是意料中的,
而且隨著輻射照度(光照)而變換。
所以光照,
如果考慮到種類的問題,
在限制和比例上都有如此大的差異,
尤其是應用在藻類的時候。
甚至在應用雷德菲爾德比例,
並且調整比例上下各達 5-10 倍的差距時,
也擔保藻類不會受到任何一方營養的限制。
Anderson 建議,
除非碳的限制是個目標,
畢竟打氣就足以提供藻類足夠的碳元素了。
很多研究文獻支持藻類的二氧化碳補償點很低,
例如一篇針對 16 種淡水藻類的研究發現,
藻類的二氧化碳補償點介於 4.8 - 41.5 μL/L,
而大部分是在 15 至 20 μL/L 之間。
換言之,
對藻類而言,
補償點最高都小於 2 ppm,
大多數的藻類是小於 1 ppm,
有些低至 0.1 ppm 的濃度。
那麼水草的二氧化碳補償點呢?
另一篇針對三種侵襲性很強的水生雜草研究發現,
黑藻(Hydrilla verticillata)的二氧化碳補償點是 44 μL/L,
金魚藻(Ceratophyllum demersum)則是 41 μL/L,
而聚藻(Myriophyllum spicatum)的二氧化碳補償點最低僅為 19 μL/L。
如果和一般水草比較起來,
侵襲性很強的水生雜草之二氧化碳飽和點介於 20-30 ppm,
這些水草具備優異的二氧化碳和重碳酸(HCO3-)的吸收機制,
這也是為何被稱為很具競爭力和侵襲性的原因。
如果考慮到種類的問題,
在限制和比例上都有如此大的差異,
尤其是應用在藻類的時候。
甚至在應用雷德菲爾德比例,
並且調整比例上下各達 5-10 倍的差距時,
也擔保藻類不會受到任何一方營養的限制。
Anderson 建議,
除非碳的限制是個目標,
畢竟打氣就足以提供藻類足夠的碳元素了。
很多研究文獻支持藻類的二氧化碳補償點很低,
例如一篇針對 16 種淡水藻類的研究發現,
藻類的二氧化碳補償點介於 4.8 - 41.5 μL/L,
而大部分是在 15 至 20 μL/L 之間。
換言之,
對藻類而言,
補償點最高都小於 2 ppm,
大多數的藻類是小於 1 ppm,
有些低至 0.1 ppm 的濃度。
那麼水草的二氧化碳補償點呢?
另一篇針對三種侵襲性很強的水生雜草研究發現,
黑藻(Hydrilla verticillata)的二氧化碳補償點是 44 μL/L,
金魚藻(Ceratophyllum demersum)則是 41 μL/L,
而聚藻(Myriophyllum spicatum)的二氧化碳補償點最低僅為 19 μL/L。
如果和一般水草比較起來,
侵襲性很強的水生雜草之二氧化碳飽和點介於 20-30 ppm,
這些水草具備優異的二氧化碳和重碳酸(HCO3-)的吸收機制,
這也是為何被稱為很具競爭力和侵襲性的原因。
待續......

 RSS 訂閱
RSS 訂閱