
不論水草或藻類都偏好 CO2,
水中的 CO2 畢竟有限,
如何攝取其他的溶解無機碳~也就是 HCO3-,
對水草來說也是非常重要的。
水中的 CO2 畢竟有限,
如何攝取其他的溶解無機碳~也就是 HCO3-,
對水草來說也是非常重要的。
許多水草具有濃縮溶解無機碳(DIC)的機制,
藉以對抗無機碳的擴散阻力,
並且降低光呼吸作用的生理作用。
一般來說,
由於水草能夠只以核酮糖二磷酸縮化酵素(Rubisco,ribulose bis-phosphate carboxylase/oxygenase)來固定無機碳、
具有比較低的 CO2 補償點、
同時對於外在的溶解無機碳有很強的親和力,
因此水草被認為具有生物物理方面的濃縮機制。
更進一步的證據顯示,
溶解無機碳在光合作用細胞內的濃度,
超出了正常被動擴散作用下,
由細胞外 DIC 進入細胞內所能達到的濃度。
對於許多水草與海藻而言,
生物物理作用的濃縮機制是很常見的。
所以我們對於許多水草能夠利用重碳酸根(HCO3-)就不足為奇了,
尤其在 pH 高達 8-9 的水中,
重碳酸的濃度通常高出二氧化碳甚多。
研究證據顯示水草使用不同的溶解無機碳濃縮機制,
目前有三種針對 HCO3- 濃縮機制被人提出來:
包括了 HCO3- 注入質體被膜(plastid envelpe membrane)、
酸鹼區域(酸鹼極性),以及 HCO3- 在質膜(plasmalemma)上的主動運輸 。
藉以對抗無機碳的擴散阻力,
並且降低光呼吸作用的生理作用。
一般來說,
由於水草能夠只以核酮糖二磷酸縮化酵素(Rubisco,ribulose bis-phosphate carboxylase/oxygenase)來固定無機碳、
具有比較低的 CO2 補償點、
同時對於外在的溶解無機碳有很強的親和力,
因此水草被認為具有生物物理方面的濃縮機制。
更進一步的證據顯示,
溶解無機碳在光合作用細胞內的濃度,
超出了正常被動擴散作用下,
由細胞外 DIC 進入細胞內所能達到的濃度。
對於許多水草與海藻而言,
生物物理作用的濃縮機制是很常見的。
所以我們對於許多水草能夠利用重碳酸根(HCO3-)就不足為奇了,
尤其在 pH 高達 8-9 的水中,
重碳酸的濃度通常高出二氧化碳甚多。
研究證據顯示水草使用不同的溶解無機碳濃縮機制,
目前有三種針對 HCO3- 濃縮機制被人提出來:
包括了 HCO3- 注入質體被膜(plastid envelpe membrane)、
酸鹼區域(酸鹼極性),以及 HCO3- 在質膜(plasmalemma)上的主動運輸 。
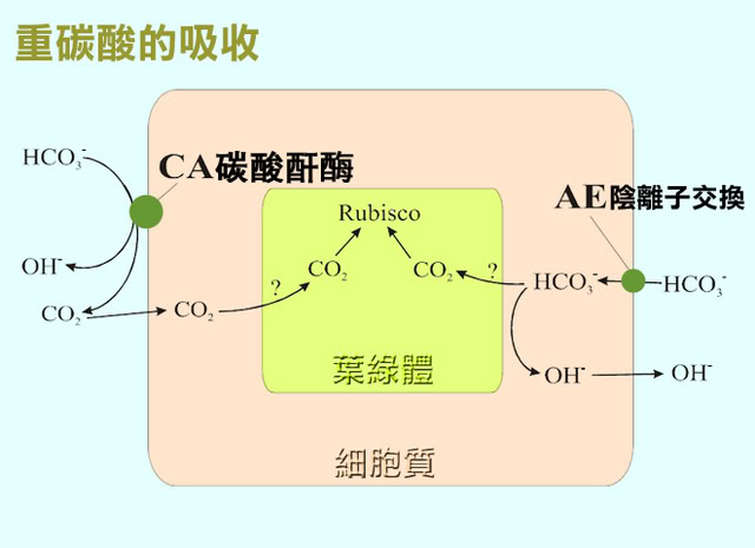
重碳酸根的主動運輸需要位於細胞內的碳酸酐酶(CA,carbonic anhydrase)的催化,
以便將 HCO3- 轉變成 CO2 好讓 rubisco 來進行碳固定。
這樣的機制在一些單細胞藻類的研究已經受到證實。
不過對於水草而言,
絕大部分的碳酸酐是作用在細胞外。
有些水草如伊樂藻(Elodea mutallii)、黑藻(Hydrilla verticillata)與光葉眼子菜(Potamogeton lucens)會在葉片的表面製造酸性與鹼性區域(酸鹼極性),
以黑藻為例,
其葉片的遠軸側(abaxial side)之 pH 為 4.0,
而葉片的近軸側(adaxial side)之 pH 為 10.5,
因此 Raven 於 1985 年的研究認為 HCO3- 從酸性區域以被動擴散作用進入植物葉片並且轉換成 CO2,
然後 CO2 又擴散至光合作用的細胞內。
這個酸性區域可以位於界線層內或者細胞壁內。
Prins 等人於 1985 年則發現光葉眼子菜(Potamogeton lucens)對於 HCO3- 的吸收是一種受化學滲透驅動的過程。
HCO3- 轉化成 CO2 位於表皮層旁邊,
受到光照的影響,
以射出氫離子(H+)的方式造成葉面上的電位差異,
並且只有 CO2 能夠通過質膜並進入葉片細胞中的細胞質。
無論如何,
Eighmy 等人於 1991 年的研究發現,
除了葉片的極性以外,
伊樂藻(Elodea mutallii)還能夠對抗濃度差異直接透過質膜來吸收 HCO3-。
此研究證實了直接吸收 HCO3- 進入細胞內,
但 HCO3- 如何在細胞內轉換成 CO2 以供 Rubisco 使用則還不清楚。
其他的水草如狐尾藻屬(Myriophyllum)與苦草屬(Vallisneria) 並沒有葉片的極性發生,
這些水草的碳酸酐酶(CA)表皮層的細胞壁或者質膜內,
碳酸酐酶催化 HCO3- 轉化成 CO2,
被認為在許多沈水植物與單細胞藻類扮演者非常重要的角色。
植物將 HCO3- 吸收進入質膜是一個必須消耗能量的過程。
無論如何,
植物利用碳酸酐酶也有其環境條件,
例如高溫、強光與非常低的 CO2/HCO3- 比值,
在這些狀況下,
直接攝取 HCO3- 是個優勢。
Axelsson 等人於 1999 年發現了有些大型的海藻除了直接由質膜吸收 HCO3- 以外,
還配合了陰離子交換(AE,anion exchange)的機制的共同作用。
類似的機轉也發生於淡水植物毛柄水毛茛(Ranunculus trichophyllus)上。
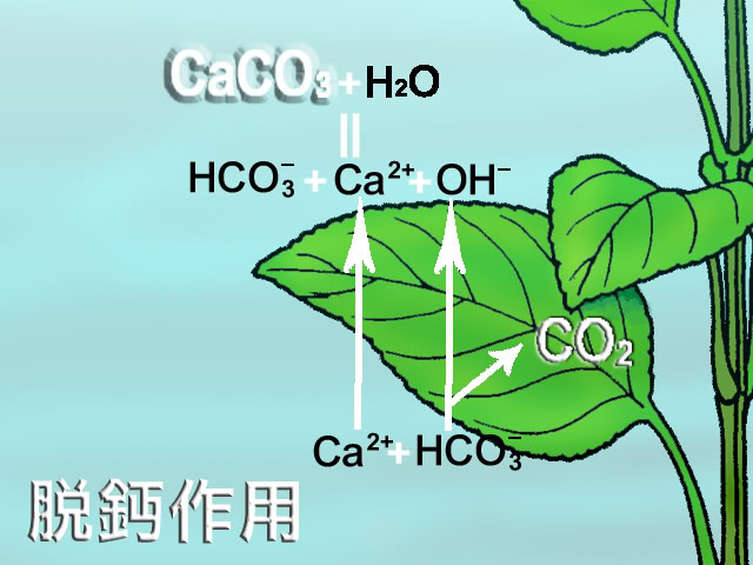
而有些水草在水中缺乏 CO2 並且吸收 HCO3- 時,
鈣離子(Ca+2)與重碳酸根(HCO3-)會同時進入葉片細胞內,
隨後 Ca2+ 又與氫氧根(OH-)一起排出至葉片細胞的表面,
鈣離子此時又與水中的重碳酸根發生化學作用,
產生了碳酸鈣(CaCO3)
Ca+2 + HCO3- + OH- -> CaCO3 + H2O
這就是我們所熟知的脫鈣作用。
本文同時刊載於台灣「AquaPets」雙月刊 2008 年八月號。

 RSS 訂閱
RSS 訂閱