
陸生植物對於碳元素的吸收,
直接來自 CO2 即可;
但對於水草而言,
必須發展出另外的機制來吸收其它的溶解無機碳(DIC, dissolved inorganic carbon),
單靠 CO2 恐怕無法順利在水底世界中生存。
直接來自 CO2 即可;
但對於水草而言,
必須發展出另外的機制來吸收其它的溶解無機碳(DIC, dissolved inorganic carbon),
單靠 CO2 恐怕無法順利在水底世界中生存。
溶解無機碳(DIC)包括了溶解於水中的二氧化碳(CO2)、重碳酸根離子(HCO3-)與碳酸根離子(CO3=),
溶解於水中的二氧化碳,
如果大氣平衡的時候,
濃度約為 0.01 mM(即 0.44 ppm),
這個濃度對陸生植物而言是很足夠的。
然而對於淡水環境而言,
溶解於水中的 CO2 變化很大,
這不只是因為環境的關係,
也會因為時間而變化。
溶解於水中的二氧化碳,
如果大氣平衡的時候,
濃度約為 0.01 mM(即 0.44 ppm),
這個濃度對陸生植物而言是很足夠的。
然而對於淡水環境而言,
溶解於水中的 CO2 變化很大,
這不只是因為環境的關係,
也會因為時間而變化。
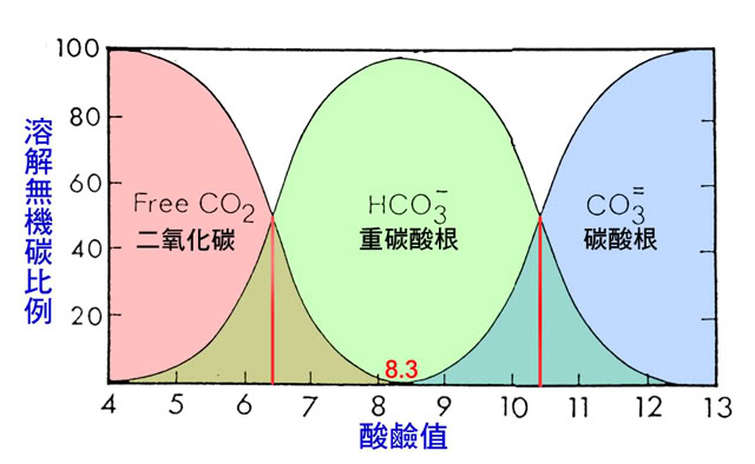
大部分的淡水水域之 pH 值位於 7-8.5 之間,
其典型的 HCO3-:CO2 倍數介於 4 至 140 倍。
相對而言,
海水環境中的變動就比較小,
況且有更高且穩定的 HCO3- 濃度(2 mM)。
不論如何,
重碳酸根離子的濃度都比溶解的二氧化碳來得高,
我們不難預料水草會吸收 HCO3- 來作為光合作用的主要碳元素來源。
許多水草都能利用 HCO3-,
除非是來自水質非常軟的環境。
因此水草無法利用 HCO3- 作為碳元素來源。
就算是能夠使用 HCO3- 作為碳元素來源,
水草還是在富含溶解 CO2 的環境中生長比較快速。
但也有例外的狀況,
例如黑藻(Hydrilla verticllata)在 pH 值為 9 的環境下,
其光合作用的速率高達 10 倍,
在這樣的 pH 值(pH > 8.3)中,
水中的溶解無機碳幾乎全部都是 HCO3-!
而水草雖然都偏好 CO2,
但在 pH < 4 的環境中,
雖然水中含有非常豐富的溶解 CO2,
但大部分的水草卻無法在此環境中生存,
這當然也與其他營養的吸收有所關連。
在淡水的環境中,
兼性(facultative)重碳酸使用者比較常見的可能原因是,
吸收 HCO3- 是需要消耗能量的,
這在很多環境條件下未必是個好處。
在海水的環境中,
由於高濃度且穩定的 HCO3-,
我們能夠預期專性(obligate)使用重碳酸的海草會比較常見。
然而海草對於 HCO3- 的吸收是否會受到水中自由態 CO2 的影響,
我們不得而知。
而許多水草雖然能夠使用 HCO3-,
但到底水草是直接固定碳酸根離子(CO3=),
或者先將 HCO3- 轉化成 CO2 後再利用,
還不是非常的清楚。
雖然生存環境有很大的差異性,
但沈水植物與陸生植物都有共同的特徵:
二氧化碳必須先溶於水中才得以進入光合作用的細胞。
可是沈水植物所需要的擴散路徑,
與陸生植物比較起來是非常漫長的;
況且 CO2 在水中的擴散速率只有在空氣中之一萬分之一。
溶解無機碳與氧氣必須進入水草葉面表皮的距離,
稱之為界線層(boundary layer)。
界線層越厚,
擴散的阻力就越大。
根據 Hutchinson 於 1975 年的研究,
擬金髮蘚(Polytrichum formosum)在水中需要相對於空氣中 10 倍的 CO2 濃度,
以達到相同的光合作用速率。
更雪上加霜的是,
水草表面所附生的藻類,
不但掠奪了水草所需的 CO2,
同時還更進一步增加了擴散的距離或界線層的厚度。
其典型的 HCO3-:CO2 倍數介於 4 至 140 倍。
相對而言,
海水環境中的變動就比較小,
況且有更高且穩定的 HCO3- 濃度(2 mM)。
不論如何,
重碳酸根離子的濃度都比溶解的二氧化碳來得高,
我們不難預料水草會吸收 HCO3- 來作為光合作用的主要碳元素來源。
許多水草都能利用 HCO3-,
除非是來自水質非常軟的環境。
因此水草無法利用 HCO3- 作為碳元素來源。
就算是能夠使用 HCO3- 作為碳元素來源,
水草還是在富含溶解 CO2 的環境中生長比較快速。
但也有例外的狀況,
例如黑藻(Hydrilla verticllata)在 pH 值為 9 的環境下,
其光合作用的速率高達 10 倍,
在這樣的 pH 值(pH > 8.3)中,
水中的溶解無機碳幾乎全部都是 HCO3-!
而水草雖然都偏好 CO2,
但在 pH < 4 的環境中,
雖然水中含有非常豐富的溶解 CO2,
但大部分的水草卻無法在此環境中生存,
這當然也與其他營養的吸收有所關連。
在淡水的環境中,
兼性(facultative)重碳酸使用者比較常見的可能原因是,
吸收 HCO3- 是需要消耗能量的,
這在很多環境條件下未必是個好處。
在海水的環境中,
由於高濃度且穩定的 HCO3-,
我們能夠預期專性(obligate)使用重碳酸的海草會比較常見。
然而海草對於 HCO3- 的吸收是否會受到水中自由態 CO2 的影響,
我們不得而知。
而許多水草雖然能夠使用 HCO3-,
但到底水草是直接固定碳酸根離子(CO3=),
或者先將 HCO3- 轉化成 CO2 後再利用,
還不是非常的清楚。
雖然生存環境有很大的差異性,
但沈水植物與陸生植物都有共同的特徵:
二氧化碳必須先溶於水中才得以進入光合作用的細胞。
可是沈水植物所需要的擴散路徑,
與陸生植物比較起來是非常漫長的;
況且 CO2 在水中的擴散速率只有在空氣中之一萬分之一。
溶解無機碳與氧氣必須進入水草葉面表皮的距離,
稱之為界線層(boundary layer)。
界線層越厚,
擴散的阻力就越大。
根據 Hutchinson 於 1975 年的研究,
擬金髮蘚(Polytrichum formosum)在水中需要相對於空氣中 10 倍的 CO2 濃度,
以達到相同的光合作用速率。
更雪上加霜的是,
水草表面所附生的藻類,
不但掠奪了水草所需的 CO2,
同時還更進一步增加了擴散的距離或界線層的厚度。
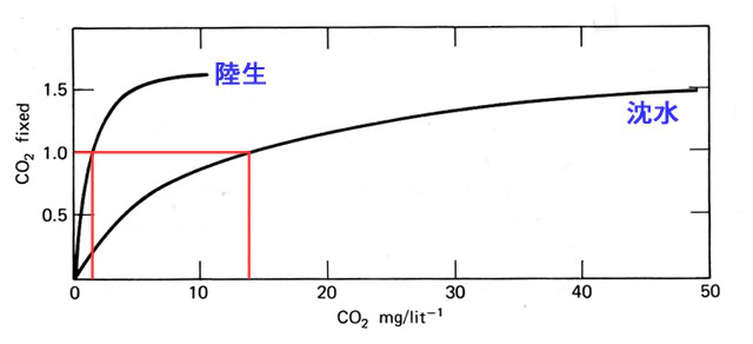
很多草友們在追求 CO2 的溶解度之餘,
假如瞭解到液態的 CO2 不但擴散速率很慢,
況且在水中的 CO2 還會轉變成 HCO3- 甚至 CO3= 等水草不易吸收的形式,
或許也該考慮使用 Tom Barr 提出的 CO2 霧氣論點來增進水草的光合作用了。
草友們可以參考一下「CO2 溶解不完全?」一文。
假如瞭解到液態的 CO2 不但擴散速率很慢,
況且在水中的 CO2 還會轉變成 HCO3- 甚至 CO3= 等水草不易吸收的形式,
或許也該考慮使用 Tom Barr 提出的 CO2 霧氣論點來增進水草的光合作用了。
草友們可以參考一下「CO2 溶解不完全?」一文。

 RSS 訂閱
RSS 訂閱